|
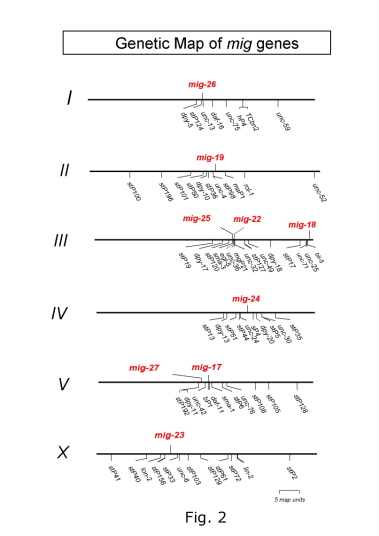
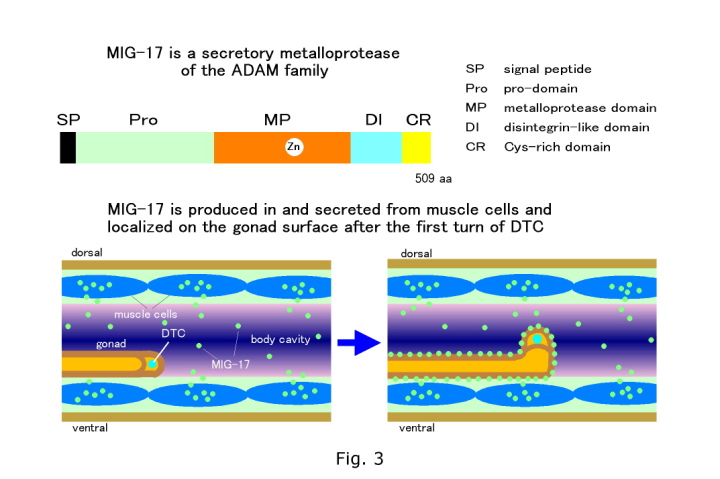
動物の発生では器官は基底膜に包まれた小さな原基から正確な細胞分裂、細胞移動および細胞形態の変化により形成される。中でも細胞移動は器官形態形成においてきわめて重要な過程である。線虫 C. elegans では受精卵から成虫に至る細胞系譜が完全に解明されており、発生過程での細胞移動も個々の細胞単位で全て記述されている。 C. elegans は細胞生物学、遺伝学および分子生物学的解析がもっとも簡単に行えるモデル動物である。我々は C. elegans 雌雄同体の生殖巣の前後両端に位置する遠端細胞(distal tip cell: DTC)の移動に注目し研究を進めている(Fig. 1)。DTC は中胚葉性の細胞で幼虫期に体壁内面に沿ってU 字型の移動を行う。 DTCの移動は生殖巣管の伸長とカップルしており、生殖巣基底膜と体壁基底膜との適切な相互作用が必要である。これまで知られている DTC 移動に必要な分子は全て脊椎動物の相同分子が存在する。例えば体壁上のガイド分子である UNC-6 はネトリン、その DTC上の受容体であるUNC-40はDCC (Deleted in Colorectal Cancer)の相同分子である(Culotti and Merz, (1998) Curr. Opin. Cell Biol. 10, 609)。このようなことから DTC の移動は動物の発生過程での器官形成機構を探る優れたモデル系であると考えられる。 これまで20種類程度の分子がDTC移動に必要であることが報告されているが、DTC移動機構に関する我々の知識はまだまだ不十分である。DTC移動異常変異体の分離・解析から、我々はDTCが蛇行・迷走する新しい表現型を示す変異体を見出した(Fig. 1) (Nishiwaki, (1999) Genetics 152, 985)。これらの変異体はDTCの移動自体には影響がないが、移動の方向あるいは経路が異常となる。21種類の変異体は9種類の遺伝子座に分類された(Fig. 2)。種々の二重変異体の組み合わせでDTCの蛇行表現型が増強される。このことはこれらの変異が同一かあるいは並列の関係にあるプロセスに影響を与えていることを示唆する。我々の研究室ではDTCの蛇行・迷走表現型を示す変異体に注目し、DTC移動に関わるさらなる分子の同定と分子間相互作用の解析を通して、DTC移動制御の新たな側面を明らかにしたいと考えている。mig-17 はADAM (A Disintegrin And Metalloprotease)ファミリーに属する Znメタロプロテアーゼ(MIG-17と命名)をコードする (Fig. 3) (Nishiwaki et al., (2000) Science 288, 2205)。ADAMファミリーはここ10数年ほどの間に見つかってきた新しいタンパク質のグループであり、メタロプロテアーゼドメインとそれに続きディスインテグリンドメインを持つことを特徴とする。以前から知られている膜型のADAMと異なり、MIG-17は 膜貫通ドメインを持たない分泌タンパク質である。MIG-17は癌の進行に伴い発現が上昇することが知られているマウスのADAMTS-1と相同性が高い (Kuno et al., 1997 J. Biol. Chem. 272, 556)。GFP融合遺伝子による解析からMIG-17は体壁筋細胞で生産、分泌され、生殖巣の表層(おそらく基底膜上)に局在することが明らかとなった(Fig. 3)。興味深いことにMIG-17はDTCが一回目の方向転換を行った後に生殖巣上に局在し始める。この局在の時期はmig-17 変異体において最初にDTC移動異常が現れる時期に一致 している。MIG-17をDTC特異的に発現させてもDTC移動異常が回復することから、MIG-17のDTC上への局在が正常な移動に重要であることが示唆されている。MIG-17メタロプロテアーゼは生殖巣あるいは体壁基底膜中の基質(未知)を分解することにより、DTCがUNC-6などの道標分子に沿って移動することを可能にしているものと思われる。 膜型ADAMのいくつかは膜結合型のシグナル分子をプロテアーゼ活性で切断し可溶化することにより、細胞外のシグナル伝達に関与することが知られている。 たとえばmeltrin-g/ADAM9はPKCdに結合し、PKCの活性化にともない膜型のEGF様成長因子を細胞から切り離す(Izumi et al., (1998) EMBO J. 17, 7260)。 ショウジョウバエのkuzbanian/ ADAM10は膜受容体NotchのリガンドであるDeltaをプロセッシングすることが報告された(Qi et al., (1999) Science 283, 91) 。分泌型ADAMは種々の病態において重要な役割をはたすことが徐々に分かってきている。aggrecanase-1, -2/ ADAMTS-4, -5は関節炎において軟骨のプロテオグリカン aggrecanを分解することが知られている(Tortorella et al., (1999) Science 284, 1664)。また、ADAMTS13は遺伝性の血栓性血小板減少性紫斑病の原因遺伝子であり 、血漿中のvon Willebrand factorの分解に関与することが示唆されている(Levy, et al., (2001) Nature 413, 488)。このように分泌型ADAMは主として細胞外マトリクスタンパク質の分解に関与する。MIG-17は分泌型ADAMの機能とその制御系を解析できるすぐれたモデル系である。 |
|